




Роль и место проблематики биофизики воды как науки
 Современные биологические и медицинские науки охватывают широкий спектр дисциплин, направленных на решение проблем сохранения устойчивости биосферы и поддержания здоровья человека в условиях роста антропогенной нагрузки. Среди них выделяется относительно новая дисциплина как биофизика, имеющая такие направления как биофизика сложных систем, молекулярная биофизика, биофизика клетки и мембранных процессов, биофизика фотобиологических процессов, радиационная биофизика и другие.
Современные биологические и медицинские науки охватывают широкий спектр дисциплин, направленных на решение проблем сохранения устойчивости биосферы и поддержания здоровья человека в условиях роста антропогенной нагрузки. Среди них выделяется относительно новая дисциплина как биофизика, имеющая такие направления как биофизика сложных систем, молекулярная биофизика, биофизика клетки и мембранных процессов, биофизика фотобиологических процессов, радиационная биофизика и другие.
Обобщенно можно сказать, что биофизика:
- это раздел биологии, изучающий физические аспекты существования живой природы на всех её уровнях, начиная от молекул и клеток и заканчивая биосферой в целом;
- это наука о физических процессах, протекающих в биологических системах разного уровня организации, и о влиянии на биологические объекты различных физических факторов.
Подобные определения науки биофизики отражают постулат о белке как первооснове жизни, в построении которого принимают участие аминокислоты, соединённые в цепь амидными связями. В данном определении, также как и в сложившихся представлениях о биофизике как науке, не упоминается вода и создается обманчивое впечатление, что белок может существовать сам по себе, то есть без воды. В то же время обезвоженные пептиды представляют собой бесструктурную массу черного цвета и только вода придает пептидам признаки живой материи и свойства структурной упорядоченности.
Таким образом, современная биофизика занята изучением «мертвой» материи, хотя оперирует таким базовым понятиям как протоплазма, не мыслимая без воды. Однако формальное признание важности воды для биологической жизни не означает понимания ее роли в ее возникновении и поддержании.
В этом отношении показательны экспериментальные работы по действию сверхмалых доз вещества, переносу биологически-активной фармакологической информации, включая генетическую. Многочисленными экспериментами установлены факты влияния зарядового состояния воды на активизацию метаболических процессов, индукцию колебательных состояний ферментов и клеточных органелл.
Так, ион-кристаллические ассоциаты пероксид анион-радикала осуществляют управление клеточным циклом. Каковы же механизмы регуляторного действия пероксид анион-радикалов?
В настоящее время в качестве регулятора метаболической активности рассматривается молекулярная перекись водорода. Ее регуляторная роль опосредуется через электрон – донорную активность NADPH – оксидазы и вовлекает также процессы фосфорилирования, действие протеин – киназ, транскрипцию регуляторных белков, осуществляемую ДНК. Транскрипционные факторы активируются плазматической перекисью водорода через активацию ядерного фактора NF-kB, протекающую в условиях глутатион — регулируемого редокс – статуса.
Регуляторные митоген — активированные протеин – киназы (МАР – киназы) также активируются Н2О2 и приводят к модуляции генной экспрессии. Активация факторов транскрипции JAK/STAT (Janus kinase/signal transducers and activators of transcription) происходит под действием Н2О2, которая оказывает влияние на ядро клетки.
Однако регуляторная роль перекиси водорода не может быть сведена исключительно к биохимическим реакциям индукции вторичных регуляторных посредников, либо поддержанию редокс – статуса клеток.
Установленный факт запаздывания генной экспрессии в генетических процессах (Bratsun D.) не может быть объяснен с классических биохимических позиций, что требует применения в анализе регуляторных функций перекиси водорода представлений о кооперативном макроскопическом квантовом поведении фазы ассоциированной воды, в которой перекись водорода находится в анион-радикальной форме в виде ассоциатов.
В последние годы авторами проведены исследования влияния перекиси водорода на состояние воды и клеточную активность. Было показано, что в диапазоне физиологических концентраций перекись водорода сильно изменяет состояние среды: уменьшаются значения водородного показателя, редокс-потенциал увеличивается на 60мВ. При этом уменьшается электропроводимость раствора и увеличивается доля фазы ассоциированной воды, что обусловлено ростом размеров ассоциатов.
Методом люминол – геминовой хемилюминесценции в растворах перекиси водорода установлена особая концентрационная точка при концентрации 45мкг/л – точка фазового перехода, ниже которой (по концентрации) перекись водорода существует исключительно в анион-радикальной форме (рисунок 1).


Рисунок 1 — Зависимость параметров люминол–геминовой хемилюминесценции (светосумма за 100 сек экспозиции –верх, время достижения максимальной интенсивности хемилюминесценции — низ) в растворах перекиси водорода
В этой точке ассоциаты способны к фазовой неустойчивости – распаду фазы, сопровождающемуся переходом фазы ассоциированной воды в свободную воду с генерацией свободных радикалов и переконденсацией электронов на другие центры захвата.
Фазовая неустойчивость – распад фазы, сопровождающийся переходом фазы ассоциированной воды в свободную с генерацией свободных радикалов и переконденсацией электронов на другие центры захвата
Σ [НО2-(*)…ОН-(*)(Н2О)тp]q → qНО2*+ qОН*+ p·q(Н2О) +Σ2е—
Репликативная активность клеток лимфоцитов крови человека (IR) в окрестности фазового перехода испытывает сильные изменения. При этом индекс репликации клеток находится в высокой корреляционной связи (коэффициент корреляции равен 0,86) с долей фазы ассоциированной воды, представленной пероксидными ассоциатами (рисунок 2).

Рисунок 2 — Репликативная активность клеток лимфоцитов крови человека в растворах перекиси водорода (коэффициент корреляции IR и доли фазы ассоциированной воды qср (уровень Dq = 0,4…0,6% ) К корр. = 0,86)
В биологических средах в присутствии катализаторов фазовый переход четко проявляется во временной динамике процессов наработки пероксид анион-радикалов. Так, в представленной на графике кривой времени максимума люминол-геминовой хемилюминесценции выделяется пик, соответствующий достижению ассоциатами максимальных размеров.

Рисунок 3 — Динамика времени максимальной интенсивности (I макс) люминол – геминовой хемилюминесценции в присутствии в воде витамина В12
Аналогичные процессы протекают в клетках с участием комплексов цитохрома С с ассоциированной водой.
Прежде, чем перейти к рассмотрению клеточной регуляторной роли пероксидных ассоциатов необходимо кратко остановиться на некоторых моментах коллективного квантового состояния зарядов, находящихся в когерентном состоянии.
В квантовом представлении коллективное состояние электронов (макроскопический волновой пакет) может быть перенесено на иные центры зацепления электронов в виде последовательности дробных возрождений. Дробные возрождения – это часть волнового пакета, несущая полный набор состояний исходного (материнского) волнового пакета. Через формирование дробных возрождений заряды могут переноситься в пространстве и времени.
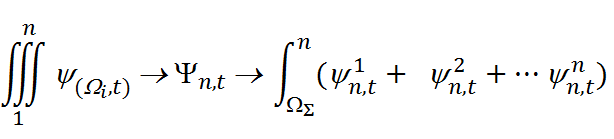
где:
![]() – делокализованное состояние в области W (область локализации макроскопического волнового пакета из n электронов);
– делокализованное состояние в области W (область локализации макроскопического волнового пакета из n электронов);
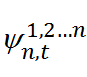 — дробные возрождения макроскопического волнового пакета на центрах пиннинга (зацепления).
— дробные возрождения макроскопического волнового пакета на центрах пиннинга (зацепления).
Волновой пакет является единым целым, рассредоточенным (делокализованном) состоянием по всей пространственно – временной области коллективного движения электронов.
Универсальное свойство дробных возрождений — функция выживания квантового макроскопического состояния электронов. Она показывает, какая часть волнового пакета вернулась в исходное положение (при распаде квантового состояния).
В результате переноса электронов (квантовой конденсации) электронов на воду или ее растворы система становится электрически неравновесной и переходит в осцилляционный режим, протекающий с периодическими изменениями своего равновесного редокс-состояния.
Рассмотренные квантовые процессы позволяют с новых позиций подойти к рассмотрению движущих сил работы митохондрий, активность которых в клеточных метаболических процессах оказывается определяющей.
Согласно квантовым представлениям движущей силой работы протонного насоса и системы окислительного фосфорилирования митохондрий являются процессы гидратации и дегидратации коферментов и железопротеинов в процессе управляемой электронами (делокализованным состоянием) кето-энольной таутомерии и квантовой конденсации электронов на парамагнитном кислороде.
Это утверждение требует кратких комментарий. Так, работами Гильберта Линга установлено, что в основе функционирования ионных насосов находятся процессы кето-энольной таутомерии полипептидных связей, регулируемой фазой ассоциированной воды. Наши исследования полностью подтверждают данные положения.
В силу неравномерного характера активации ферментов во времени их способность к конденсации электронов также осциллирует, что приводит к осцилляциям всех последующих биохимических процессов в клетке.
После того, как были рассмотрены некоторые аспекты квантового поведения ассоциатов, можно приступить к анализу их регуляторной роли в клеточном метаболизме.
Наиболее важная регуляторная роль пероксидных ассоциатов реализуется в процессе управления клеточным циклом.
В настоящее время считается, что перекись водорода принимает участие в управлении клеточным циклом, которое осуществляется путем окисления протеинкиназ. При этом имеет значение редокс-состояние среды цитоплазмы клеток. Так, митоз реализуется при значениях Еh=-240мВ, пролиферация клеток протекает при Еh до -220мВ, дифференцировка клеток – при Еh=-200мВ, а при максимальных значениях Еh=-170 мВ формируются условия для апоптоза клеток (Menon S.G. et al., 2007).
Последовательность циклов строго контролируется циклин-зависимыми протеинкиназами (cdc). К основным комплексам позвоночных относятся следующие циклины и соответствующие им протеинкиназы:
- циклин D-cdk4 (переход G0/G1);
- циклин Е -cdk2 (переход G1/S);
- циклин А-cdk2 (переход в фазу S);
- циклин В1-cdk1 (переход в фазу митоза клеток G2/M).
В цикле имеется особая точка – точка рестрикции (R), после которой клетка становится невосприимчивой к внешним сигналам вплоть до завершения всего цикла деления. В точке рестрикции клетка также может войти в апоптоз (рисунок 4).
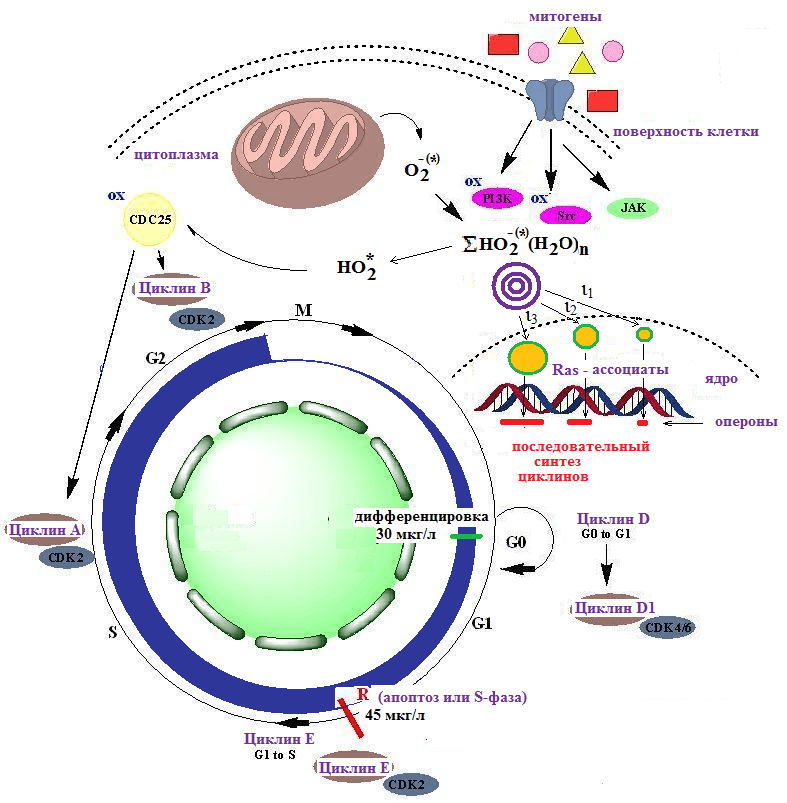
Рисунок 4 — Регулировка пероксидными ассоциатами клеточного цикла
Основное влияние ассоциатов заключается не столько в изменении редокс-состояния цитоплазмы клеток, сколько в нелокальном наведении соразмерных электростатических полей во внутриядерном пространстве, где функционируют Ras нанокластеры, выполняющие пространственно–зависимые короткодействующие сигнальные функции для активации оперонов (оперон — контролируемая единичным регуляторным сигналом или промотором функциональная единица генома ДНК, содержащая кластер активируемых генов). Внутренняя программа дифференцировки или апоптоза клеток согласно существующим представлениям связана с селективной экспрессией оперонов.
Рассматривая приведенную на слайде схему регуляторной роли пероксид анион-радикалов можно установить, что дифференцировка моноцитов в макрофаги осуществляется при достижении ассоциатами размера ~20нм. Точка рестрикции соответствует максимальному диаметру пероксид ассоциатов ~30нм. При увеличении цитозольной концентрации перекиси водорода более 45 мкг/л размеры ассоциатов существенно уменьшаются и их влияние на активацию генов сокращается.
Очевидно, что усиление активности функционирования митохондрий клеток, промотируемой активированной водой, приводит к ускорению клеточного цикла, включая дифференцировку моноцитов в макрофаги
Оценим некоторые результаты выполненных биологических исследований с применением активированной воды. Характеризуя регуляторное действие воды с электрон-донорными свойствами необходимо остановиться на некоторых критических органах и показателях.
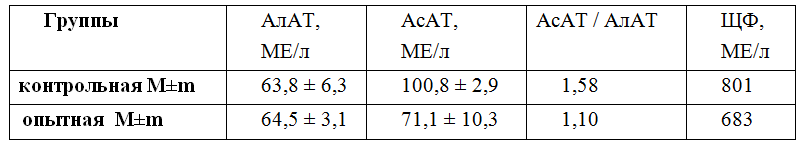
Так, при анализе функционального состояния печени среди других показателей оцениваются маркеры повреждения гепатоцитов — АсАТ (ас-паратаминотрансфераза), АлАТ (аланинаминотрансфераза), щелочная фосфатаза. Показатели АсАТ и щелочная фосфатаза при приеме активированной воды уменьшаются, обуславливая снижение значений индекса Ритиса (отношение АсАТ / АлАТ) (таблица 1).
Таблица 1 — Показатели функционального состояния печени
Активированная вода оказывает положительное влияние на морфогематологические показатели крови организма поросят. В их крови меньше нейтрофилов (на 1-11%), больше лимфоцитов (на 37,6-66,3%) при более высокой бактерицидной и лизоцимной активности сыворотки крови на фоне более низкого (на 14,3-35,9 %) содержания лейкоцитов (таблица 2).
Повышение в крови гемоглобина и уровня гематокрита (на 6,3 -11,7 % и 1,24 — 4,4 %) по отношению к контролю указывает на более активное формирование гуморального и клеточного иммунитета, а также неспецифической резистентности организма в целом.
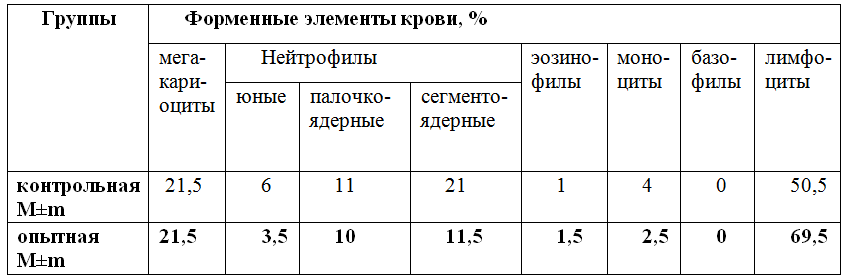
После поения свиней активированной водой изменяется макро- и микроэлементный состав жидких сред и органов животных. Уровни кальция и магния в берцовой кости достоверно увеличиваются, а в плазме крови слегка снижаются. Содержание железа в селезенке и почке существенно увеличивается, что отражает процессы их активации. Увеличивается содержание микроэлементов и фосфора. Следует отметить, что основная доля микро- и макро- элементов увеличивается вследствие лучшего усвоение из пищи (таблица 3).
Таблица 3 – изменение уровня макро- и микроэлементов в организме поросят после постоянного приема активированной воды с электрон-донорными свойствами
- Прием активированной воды с электрон-донорными свойствами стимулирует рост, развитие и предотвращает заболевания птиц и животных. Активированная вода:
- способствует интенсификации и нормализации обмена веществ;
- оказывает положительное влияние на сохранность поголовья, повышает его адаптационную способность к стрессовым условиям;
- улучшает физиолого-биохимические показатели и функциональное состояние внутренних органов, обеспечивает формирование более устойчивого клеточного иммунитета и «продуктивного здоровья» молодняка, что позволяет реализовать их генетически обусловленный потенциал;
- ускоряет костеобразование соответственно возрасту животного и сращивание переломов костей и стимулирует восстановление их прочности и нормальной структуры.
Рассмотренные выше вопросы регуляторной роли пероксидных ассоциатов и полученные подтверждения биологической активности питьевой воды с электрон-донорными свойствами позволяют оценить ее влияния на таких процессах как ремоделирование и репарация кости.
Патологический процесс остеопороза промотируется метаболическими изменениями в базисной мультиклеточной единице под влиянием гормональных изменений, инородных клеток, микроорганизмов и неблагоприятных факторов окружающей среды.
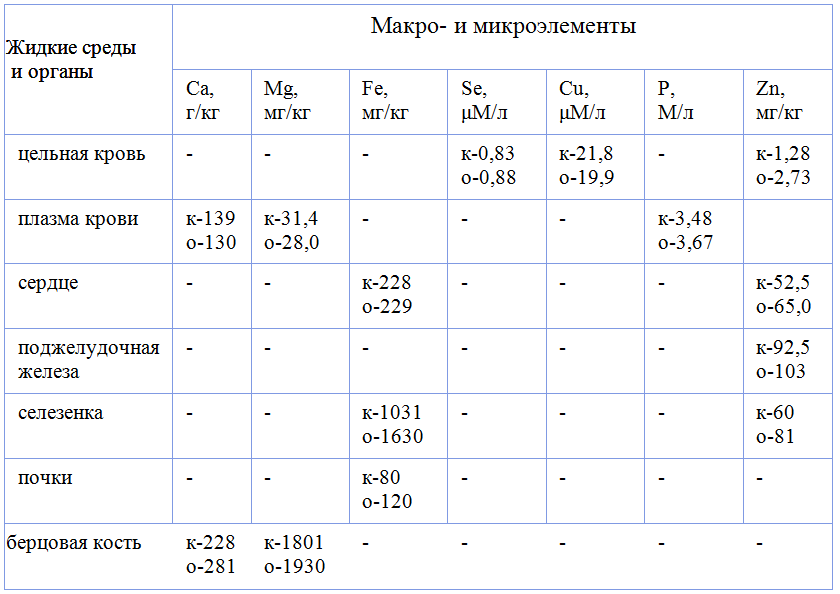
В настоящее время в качестве управляющей системы костного ремоделирования рассматривается система активации рецептора для ядерного factor κаппа В — RANK — The receptor activator for nuclear factor κВ, растворимого лиганда — RANKL — RANK kB ligand и остеопротегерин — OPG – osteoprotegerin.
На рисунке 5 приведена модель, иллюстрирующая механизм, посредством которого остеобласт/стромальные клетки регулируют дифференцировку и активацию остеокластов. Три регуляторных фактора — гидролизованная форма витамина — D3 1,25(OH)2D3, паратгормон — РТН/PGE2 и интерлейкин (IL) -11 индуцируют в остеобласт/стромальных клетках лиганды рецептора активации ядерного фактора-кВ (RANKL) .
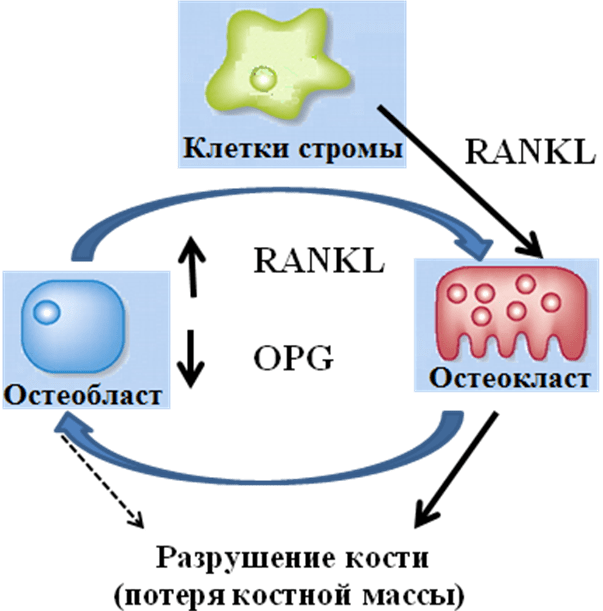
Рисунок 5 — Патогенетический путь разрушения кости
RANKL передает сигнал остеокластогенеза через RANK рецепторы предшественников остеокластов.
Остеопротегерин (ОПГ) ингибирует остеокластогенез, прерывая связывание RANKL с RANK. Макрофагальный колониестимулирующий фактор (M-CSF), полученный с помощью остеобласт/стромальных клеток, также необходим для пролиферации и дифференцировки клеток-предшественников остеокластов. Увеличение соотношения RANKL-к-OPG приводит к увеличению образования остеокластов, что увеличивает скорость ремоделирования и/или метаболизма костной ткани. Патологические изменения состояния кости характеризуется повышенной активностью остеокластов.
В обход гормональной системы существуют несколько способов влияния на активность предшественников остеокластов и регуляцию остеокластогенеза: 1- устранение источников sRANKL (таких как паразитирующие микроорганизмы и вызываемые ими воспалительные процессы, инородные клетки аденом, онкоклетки и др.), 2- ингибирование активности RANK – лиганда (например, с помощью высокоспецифичных моноклональных антител — Деносумаба , 3 — путем неспецифической активации митотической активности и процессов дифференцировки предшественников (моноцитов и макрофагов) по механизмам, не связанным с RANKL (sRANKL)/OPG – регулированием, стимулирование синтеза остеобласт/стромальными клетками и активированными лимфоцитами OPG при снижении активности sRANKL и RANKL регуляторов.
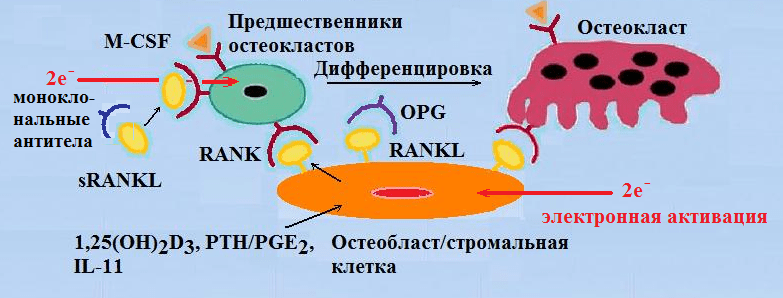
Рисунок 6 — Модель, иллюстрирующая механизм, посредством которого остеобласт/стромальные клетки регулируют дифференцировку и активацию остеокластов
Последний способ, в отличие от использования моноклональных антител, позволяет не только изменить соотношение регуляторов в сторону увеличения концентрации OPG, но и ускорить процесс остеокластогенеза, восстановить физиологически полноценную внутриклеточную среду: повысить рН, уменьшить Еh и вязкость, увеличить гидратацию белковых структур и таким образом восстановить нормальные условия для конформационной активности, электронного транспорта и транслокации внутриклеточных белковых комплексов. При этом за счет электронного стимулирования ферментативной активности достигаются эффекты ускорения митоза и пролиферации клеток – предшественников остеокластов. Одновременно увеличивается активность паратгормона и цитокинов, также принимающих участие в метаболических процессах костного ремоделирования.
Изменения состояния системы RANKL (sRANKL)/OPG можно проследить на примере лечения парадонта. Так, влияние электронной активации тканей парадонта на активность ферментов и содержание регуляторов остеогенеза приводит к снижению активности лактатдегидрогеназы (ЛДГ), аспартатаминотрансферазы (АСТ), аланинаминотрансферазы (АЛТ) и щелочной фосфатазы (ЩФ). Одновременно изменяется соотношение sRANKL)/OPG в сторону увеличения концентрации блокаторов активности остеокластов.
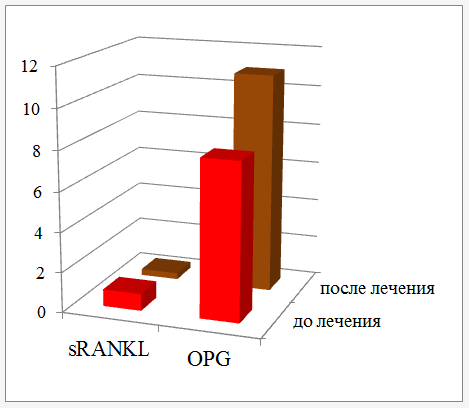
Рисунок 6 — Влияние электронной активации тканей парадонта на активность ферментов и содержание регуляторов остеогенеза
Соотношение OPG и sRANKL является информативным показателем деструкции костной ткани, что позволяет диагностировать риск развития прогрессирующего пародонтита. Электронная активация, достигаемая при применении активированной воды, изменяет соотношение регуляторов остеокластогенеза в сторону их сбалансированности.
При применении активированной воды достигаются системные эффекты. Повышение концентрации кальция и микроэлементов вследствие их лучшего усвоения из пищи в кровотоке вызывает системные эффекты (благодаря регуляторному действию кальция и микроэлементов многих биохимических процессов): поглощение кальция скелетными мышцами увеличивает их сократимость, что улучшает трофику мягких тканей травмированной конечности, а, следовательно, и способствует восстановлению функции скелетных мышц поврежденного сегмента (снижение активности креатинкиназы и лактатдегидрогеназы – ферментов маркеров скелетных мышц);
Восстановление электронного дефицита основной энергообеспечивающей клетки органеллы – митохондрии приводит к восстановлению активности дыхательного фермента цитохрома С и ко-фермента НАД, активации гликолиза и цикла трикарбоновых кислот, а также сопряженного с ними окислительного фосфорилирования. Одновременно увеличивается общее содержание адениловых нуклеотидов, что позволяет обновить и активировать пентозофосфатный путь синтеза АТФ, НАДФ и рибозы.
Антигипоксическое и противовоспалительное действие, обусловленное электронной активаций митохондриальной активности клеток, стабилизирует деструктивные процессы в костной ткани, стимулирует регенерацию эпителиальной ткани и улучшает периферическую микроциркуляцию.
При лечении водой с электрон-донорными свойствами достигаются следующие физиолого-биохимические показатели:
- оказывает положительное влияние на течение репаративных процессов в поврежденных тканях при лечении переломов кости средней и тяжелой степени тяжести.
- ее лечебное действие обусловлено антигипоксическими и детоксицирующими свойствами, способностью повышать пролиферативную активность и уровень внутриклеточного свободного кальция в остеобластах, активацией макрофагальной реакции, а также процессов неоангиогенеза и восстановления миелоидной ткани в области отломков кости после периода циркуляторной гипоксии.
- электрон-донорное действие воды приводит к снижению редокс-состояния и стабилизации рН клеток, интенсифицирует обмен веществ, включая обмен микро- и макро – элементов, способствует существенному улучшению функционирования системы антиоксидантной защиты организма, положительно влияет на физиолого-биохимические показатели и функциональное состояние внутренних органов.
- применение активированной воды способствует ускорению костеобразования, не сопровождается значительным увеличением уровня кальция в крови выше критического, восстанавливает их прочность и нормальную структуру.
Рассматривая механизмы действия активированной воды на внутриклеточные метаболические процессы следует остановиться на том, что они носят неконтактный и нелокальный характер, что не противоречит квантовым представлениям, а наоборот, является логических следствием макроскопических квантовых взаимодействий.
Высказываем рабочую гипотезу о том, что управляющее действие ассоциированной воды, обусловленное ее концентрационными и редокс- фазовыми переходами, проявляется на всех уровнях организации живого. Реализация управляющего действия фазы ассоциированной воды осуществляется в процессах резкого усиления параметров ассоциации воды, сопровождаемых индукцией нелокальной электродвижущей силы, изменениями электрохимического состояния водной среды, в первую очередь, среды клеточных органелл. Этот подход может служить логическим продолжением как известных механизмов управления колебательной активностью органелл клеток, так и позволяет установить механизмы направленного движения органелл и везикул в цитоплазме клетки.
Можно утверждать, что отмеченные выше экспериментальные данные о воде закладывают базис новой науки – биофизики воды, фундаментом которой являются представления о воде как кооперативно – организованной квантовой системе, носителем взаимодействия в которой являются делокализованные электроны в составе фазы ассоциированной воды.
Отметим еще несколько важных для биофизики воды направлений исследований. Среди них трансмутация элементов, протекающая в живом организме в результате протонных захватов. Это направление исследований развивалось в 40-х годах прошлого столетия. Анализируя биологические эффекты явления трансмутации, следует отметить и такое явление как захват атомом протона вблизи ядра (без ядерного превращения), которое также приводит к изменениям электронной конфигурации атомов.
По нашим представлениям явления захвата протонов атомами и трансмутация элементов живой ткани являются основными причинами старения организма, в связи с чем одним из направлений биофизики воды следует считать изучение процессов деградации макроскопического квантового состояния ассоциированной воды в живых организмах вследствие нарушений изотопного состава и внутриатомной структуры элементов белковых структур и воды.
Таким образом, практические приложения науки биофизики воды направлены на профилактику и лечение метаболически обусловленных заболеваний человека и поддержание жизнеспособности биоты во всех своих формах, управления биоэнергетической активностью окружающей среды, разработку теории и практики восстановления биоструктур, в том числе продления жизни.
Авторы:
© Анатолий Стехин, к.т.н.
© Ing. Mgr. Igor Oberemko
© Tinowa Group
10.03.2016